




Las células natural killer (NK) juegan un papel importante en la defensa contra las células tumorales. El desarrollo y la función de las células NK se rige por un equilibrio dinámico entre la inhibición y la activación de los receptores de la superficie celular, incluidos los receptores KIR.
Pacientes y métodose realiza un estudio de casos y controles que compara a un grupo de 46 niños diagnosticados de enfermedades malignas, el grupo control está constituido por 82 niños sanos. Se determinaron y compararon entre grupos los genes, haplotipos y ligandos KIRs.
Resultadosno existen diferencias en genes KIRs, haplotipos KIRs ni en ligandos de genes KIRs entre grupos. Sin embargo, al estudiar conjuntamente KIRs y ligandos, k2DS1_C2 fue significativamente superior en el grupo de niños oncológicos (p̊=̊0,016).
ConclusionesNuestros resultados no proporcionan evidencia de una asociación entre enfermedades oncológicas pediátricas con genotipos y grupos de genes KIRs. El genotipo k2DS1_C2 podría predisponer a la susceptibilidad a procesos malignos en la población infantil.
Natural killer (NK) cells play an important role in defense against tumor cells. The development and function of NK cells is governed by a dynamic balance between inhibition and activation of cell surface receptors, including KIR receptors.
Patients and methodA case-control study is carried out that compares a group of 46 children diagnosed with malignant diseases, the control group is made up of 82 healthy children. KIRs genes, haplotypes and ligands were determined and compared between groups.
ResultsThere are no differences in KIRs genes, KIRs haplotypes or in KIRs gene ligands between groups. However, when KIRS and ligands were jointly studied, k2DS1_C2 was significantly higher in the group of cancer children (p̊=̊0.016).
ConclusionsOur results do not provide evidence of an association between pediatric cancer disease with genotypes and groups of genes KIRs. The k2DS1_C2 genotype could predispose to susceptibility to malignant processes in children.
El desarrollo de la biología molecular ha contribuido a completar el conocimiento de la influencia de los factores genéticos e inmunes en la patogénesis del cáncer en la edad pediátrica1. Las células NK, que son componentes esenciales del sistema inmune innato, se han convertido en uno de los focos de la investigación2.
Las células natural killer (NK) actúan dirigiéndose espontáneamente contra las células consideradas peligrosas para el huésped (cáncer, células extrañas o infectadas por virus) y, por lo tanto, se presume que son efectores clave en la inmunovigilancia del cáncer, el rechazo de trasplantes y la inmunidad viral temprana3.
El desarrollo y la función de las células NK se rige por un equilibrio dinámico entre la inhibición y la activación de los receptores de la superficie celular, incluidos los receptores de KIR4,5. Los genes KIR se encuentran en el cromosoma 19q13.4. Hasta la fecha, la familia de genes KIR compromete 16 loci, incluidos 2 seudogenes (KIR2DP1 y KIR3DP1) y 4 genes KIR marco (KIR2DL4, KIR3DL2, KIR3DL3 y KIR3DP1). Catorce genes KIR, funcionales y altamente homólogos, codifican los receptores clave que desencadenan la activación (2DS1-5 y 3DS1), la inhibición (2DL1-3, 2DL5 y 3DL1-3) o la activación e inhibición (2DL4) de las células NK5.Varios estudios han analizado una posible asociación entre los genes KIR y el desarrollo de ciertas formas de cáncer infantil, fundamentalmente de leucemias6-12.
Sobre la base del contenido del gen KIR, se han reconocido grupos de dos haplotipos KIR, A y B. Los haplotipos KIR del grupo A se encuentran en todas las poblaciones y se componen de cinco KIR inhibidores (KIR2DL3, KIR2DL1, KIR2DL4, KIR3DL1, KIR3DL2) y el estimulante KIR2DS4. Los haplotipos KIR del grupo B consisten en contenido genético diverso que incluye varios genes (KIR2DL2, KIR2DL5, KIR2DS1, KIR2DS2, KIR2DS3, KIR2DS5 y KIR3DS1) que no se encuentran en los haplotipos KIR del grupo A. La mayoría de los KIRs que componen el haplotipo B poseen función activadora. Se ha informado de la incidencia de haplotipos KIR o genes KIR específicos con susceptibilidad a enfermedades humanas, como son los trastornos autoinmunes13,14, abortos involuntarios recurrentes15,16, enfermedades infecciosas17 y cánceres18.
Los genes KIR son altamente polimórficos e interactúan con la molécula clase I del antígeno leucocitario humano (HLA), igualmente polimórfico. Los genes KIR se expresan en la superficie de las células NK y algunas células T, y regulan el desarrollo y la función de estas células a través de la interacción con sus ligandos HLA afines19, de esta manera producen un efecto variado sobre la actividad de las células NK. La expresión de alelos HLA de clase I en la superficie de las células permite que las células NK se reconozcan como propias y les ayuda a dirigirse a las entidades no propias, como las células cancerosas y algunas células infectadas por virus6,20-22.
En un esfuerzo por abordar los factores genéticos que podrían contribuir a la susceptibilidad a cáncer infantil, realizamos un estudio de casos y controles y evaluamos los genes/genotipos KIR/ligandos HLA con el objetivo de investigar la asociación entre los genes/genotipos KIR en niños oncológicos
Material y métodoRegistramos 46 casos pediátricos de edades comprendidas entre 0 y 14 años, diagnosticados y tratados de enfermedades malignas entre diciembre de 2010 y marzo de 2017. Todos los casos fueron reclutados en la unidad de hematología-oncología pediátrica del Hospital Universitario Materno Infantil Virgen de las Nieves, de Granada.
Se utilizaron como controles un total de 82 niños sin antecedentes de malignidad o cualquier otra enfermedad, de edades comprendidas entre 0 y 14 años.
Se tomaron dos muestras de sangre de cada paciente, y se almacenaron alícuotas de suero y sangre total a -20̊°C y se remitieron a la red de Biobanco, donde se procesaron las muestras. Las determinaciones genéticas fueron realizadas en el Instituto de Investigación Biosanitaria de Granada.
1. Tipificación HLA: en las 128 muestras recogidas se analizan los loci HLA-A*, B* y CW* de clase I, HLA-DRB1*, DQB1* *, DQA1 y DP de clase II.
Para el genotipado se utiliza el kit LABTypeTM SSO (One Lambda, Inc. Canoga Park CA) que utiliza la tecnología Lumine xTM x MAPTM. Luminex es un sistema de reacción en cadena de polimerasa reversa de secuencias especificas con oligonucleótidos (PCR-SSO) que implica la amplificación de regiones específicas dentro del CMH I o II con grupos de cebados (primers) específicos, seguidos por un proceso de sondeo del ADN amplificado con esferas de Luminex cubiertas con probes de oligonucleótidos específicos de secuencia para identificar la presencia o ausencia de alelos específicos. La asignación del tipo HLA se basa en la reacción patrón observada y se compara con diferentes patrones conocidos.
2. Tipaje KIR: se utilizó el kit de genotipado LABType SSO® (Oligonucleótidos de secuencia específica) para 14 genes y 2 seudogenes KIR. De éstos, 8 genes tienen función inhibidora (2DL1, 2DL2, 2DL3, 2DL4, 2DL5, 3DL1, 3DL2 y 3DL3) y 6 activadora (2DS1, 2DS2, 2DS3, 2DS4, 2DS5 y 3DS1). Para la hibridación se utilizaron sondas de oligonucleótidos y amplificación por PCR de los exones 3, 4, 5, 7, 8 y 9 del cromosoma 19.
Se obtuvieron datos cuantitativos mediante el sistema Luminex-200 (Luminex Corporation, Austin, TX), y el análisis de éstos se realizó mediante el software v2.3, Luminex 100®, según las instrucciones del fabricante.
Para el estudio del haplotipo se consideró que: el paciente con haplotipo KIR del grupo A, lleva un conjunto fijo de nueve genes (KIR 3DL3, 2DL3, 2DP1, 2DL1, 3DP1, 2DL4, 3DL1, 2DS4, 2DL2) que incluyen un solo gen activador (2DS4), dos seudogenes (2DP1 y 3DP1) y seis genes inhibidores, por lo tanto, son haplotipo inhibidor. Los portadores de dos copias de los haplotipos KIR del grupo A se consideraron genotipos AA, mientras tanto, las personas que carecen de cualquiera de los cuatro genes variables (2DL1, 2DL3, 3DL1 y 2DS4) se consideraron portadores de dos copias haplotipos del grupo B (genotipos BB). Todas las combinaciones restantes se designaron como heterocigotos que portaban ambos haplogrupos, es decir, genotipos AB. Los haplotipos Bx se han caracterizado por la presencia de genes activadores adicionales y la ausencia de genes KIR inhibidores variables específicos del grupo A (2DL2, 2DL3 y 3DL1).
Análisis estadísticoEn cuanto a las variables del estudio, la variable independiente fue la presencia o ausencia de proceso maligno, las variables dependientes fueron el genotipo HLA y el tipaje KIR.
La gestión y análisis de los datos se realizó con el programa SPSS19.0 para Windows (SPSS Inc., Chicago, IL). La comparación entre grupos se realizó mediante la prueba de chi-cuadrado para las variables cualitativas y t de Student para variables cuantitativas.
El efecto independiente de cada factor se determinó usando un modelo de regresión logística.
Se consideró que p < 0,05 era estadísticamente significativo.
La gestión y análisis de los datos se realizó con el programa SPSS 19.0 para Windows (SPSS Inc., Chicago, IL).
Declaración de éticaEl presente estudio se ha llevado a cabo de acuerdo con la última versión de la Declaración de Helsinki. Todos los casos y controles han firmado el consentimiento informado. El proyecto cuenta con la aprobación del Comité de Ética Provincial.
ResultadosEl grupo de pacientes oncológicos estuvo constituido por un total de 29 hombres (63%) y 17 mujeres (37%) de edades comprendidas entre 0 y 14 años. En cuanto a la enfermedad oncológica, se trató de 38 leucemias linfoblásticas agudas, cuatro linfomas linfoblásticos y cuatro histiocitosis. De los 82 niños que conformaron el grupo control, 48 fueron hombres (58,54%) y 34 mujeres (41,46%).
KIRsEn la figura 1 se presenta la distribución de KIRs en los pacientes oncológicos y en el grupo control.

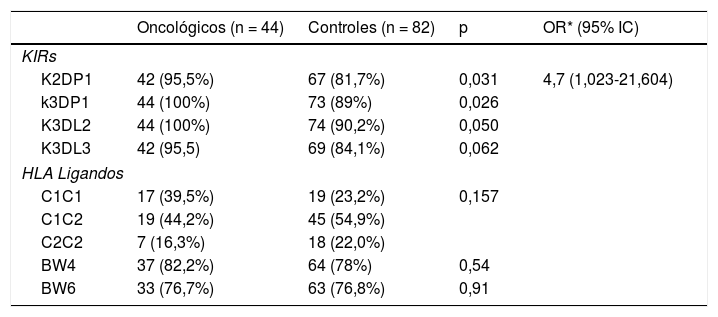
Se encontraron diferencias significativas en los KIRS (seudogenes) K2DP1, K3DP1. Los KIRs inhibidores k3DL2 y k3DL3 se encontraron en el límite de la significación (tabla 1).
Comparaciones entre grupo de niños oncológico y grupo control de KIRs y HLA ligandos
| Oncológicos (n = 44) | Controles (n = 82) | p | OR* (95% IC) | |
|---|---|---|---|---|
| KIRs | ||||
| K2DP1 | 42 (95,5%) | 67 (81,7%) | 0,031 | 4,7 (1,023-21,604) |
| k3DP1 | 44 (100%) | 73 (89%) | 0,026 | |
| K3DL2 | 44 (100%) | 74 (90,2%) | 0,050 | |
| K3DL3 | 42 (95,5) | 69 (84,1%) | 0,062 | |
| HLA Ligandos | ||||
| C1C1 | 17 (39,5%) | 19 (23,2%) | 0,157 | |
| C1C2 | 19 (44,2%) | 45 (54,9%) | ||
| C2C2 | 7 (16,3%) | 18 (22,0%) | ||
| BW4 | 37 (82,2%) | 64 (78%) | 0,54 | |
| BW6 | 33 (76,7%) | 63 (76,8%) | 0,91 | |
A pesar de haber niños que tienen presentes todos los KIRs activadores y otros niños con todos los KIRs inhibidores, al comparar ambos grupos con respecto a este parámetro, no se encontraron diferencias significativas.
Haplotipos KIRsAl comparar los haplotipos KIRs no se observaron diferencias entre grupos (fig. 2).
Ligandos KIRs
Se procedió, posteriormente, al estudio de los ligandos KIRs (C1, C2, Bw4 y Bw6). El estudio entre grupos no mostró significación estadística (tabla 1).
KIRs-ligandosk2DS1_C2L fue significativamente superior en el grupo de enfermos oncológicos (p̊=̊0,016). Sin embargo, la OR de 0,489 (0,224-1,069) con 1 grado de libertad, perdió la significación estadística (p̊=̊0,07).
Otros estudios KIRsPuesto que los alelos KIR2DL2 y KIR 2DL3 son alelos de un mismo gen, se planteó realizar comparación entre: 2DL2/2DL2 (homocigótica para 2DL2), 2DL3/2DL3 (homocigótica para 2DL3), y 2DL2/2DL3 (heterocigótica). No se encontraron diferencias significativas al realizar esta comparación (fig. 3). De igual forma, puesto que los alelos KIR3DL1 y KIR3DS1 son alelos de un mismo gen, el estudio se realizó comparando: 3DL1/3DL1 (homocigótica para 3DL1), 3DS1/3DS1 (homocigótica para 3DS1), y 3DL1/3DS1 (heterocigótica). No se obtuvieron diferencias significativas entre grupos (fig. 4).
Discusión

El repertorio de genes KIR reconoce locus específicos de moléculas de HLA de clase I y forma una serie de interacciones receptor-ligando que determina la respuesta de las células NK5. Las interacciones de las células NK dependen de las combinaciones de productos génicos variables KIR y HLA de clase I. La activación de las células NK depende del equilibrio entre los receptores inhibidores y activadores. En este estudio, se realizó un análisis de la presencia o ausencia de genes de 16 KIR, incluyendo tanto KIR inhibidores como activadores. También se ha realizado el análisis de los KIRs y sus ligandos en niños con enfermedades malignas comparado con un grupo control y el análisis de haplotipos KIRs.
Debido al papel crucial de los receptores KIR en la regulación de la actividad de las células NK, se han realizado análisis de genotipos KIR en diversas afecciones, oncológica en niños6,7,9,10,23,24 para dilucidar la contribución potencial de la variabilidad de KIR en la susceptibilidad a la enfermedad y la respuesta de la misma al tratamiento.
Se observó que ninguno de los genes KIR individuales (ni activadores ni inhibidores), tendía a aumentar o reducir el riesgo de desarrollar enfermedades malignas en la población estudiada. Sólo se encontraron diferencias significativas en los KIRS (seudogenes) K2DP1, K3DP1. La falta de asociación de los KIRs individuales con las enfermedades malignas no está correctamente aclarada. Estudios previos arrojan resultados contradictorios en este sentido: Almalte et al.25 realizaron un estudio de casos y controles en pacientes pediátricos canadienses de origen francés mediante el estudio de la frecuencia de los 6 genes KIR activadores encontrando que portar genes KIR activadores se asocia con un riesgo menor de desarrollar leucemia linfoblástica aguda de células B (LLA-B) en estos niños, y cuanto mayor es el número de genes KIR activadores, menor es el riesgo de desarrollar la enfermedad.
Misra et al.9 apoyan la opinión de los efectos acumulativos de la activación e inhibición de señales sobre la modulación de la función efectora de las células NK en los casos de LLA infantil. En este estudio se encontró un cambio hacia la sobreactivación de las células NK en función del contenido del gen KIR del individuo, que comprende un mayor número de genes activadores y un número reducido de inhibidores (KIR2DL3, KIR2DL2 y KIR2DL5) en los casos de LLA infantil en comparación con los controles.
Por el contrario, Babor et al.8 no informaron asociación entre los genes KIR individuales y la LLA-B infantil en niños de origen europeo. Tampoco se encontró asociación en el estudio de Jiang et al.6.
En el presente trabajo ninguno de los haplotipos KIRs se correlacionó con el riesgo de enfermedad maligna en la infancia. de Smith et al.26 encontraron que la frecuencia del haplotipo A/A era más frecuente en niños con LLA, este hallazgo se encontró también en el estudio de Almalte et al.25. Un estudio27 mostró que ser homocigótico A/A ofrecía protección contra la leucemia en adultos en el sur de China. Las células NK de los individuos homocigotos para el KIR A fueron fuertemente citotóxicas para las células leucémicas y la incidencia de activación de los KIR aumentó en los casos de LLA infantil. En el estudio de Misra et al.9 las personas que tenían genotipos BB tenían un mayor riesgo de LLA infantil (2,5 veces) que las que tenían genotipos AA o AB.
Para combinaciones de genes KIR inhibidores y activadores específicos y sus ligandos HLA, el presente trabajo pone de manifiesto un riesgo aumentado de enfermedad maligna en la combinación K2DS1-HLAC2. Este riesgo ha sido también puesto de manifiesto anteriormente9 y puede deberse al desencadenamiento de una hiperreactividad localizada inadecuada de las células NK que exacerbe el riesgo de crecimiento de células leucémicas. A pesar de que se desconoce el significado exacto de este hallazgo, dado que pueden heredarse diferentes números de KIR activadores entre individuos, las frecuencias de varias combinaciones de KIR activadores y sus ligandos HLA de clase I afines podrían modular el riesgo de tumores en la infancia. Los informes contradictorios pueden estar influenciados por las diferentes etnias y diferentes tumores analizados.
A la vista de los presentes resultados, recomendamos una mayor investigación de la expresión y función de los genes KIR, ligandos HLA y la expresión de moléculas HLA clase I en células tumorales para comprender mejor los mecanismos subyacentes de la posible disfunción de las células NK en el cáncer.
FinanciaciónEl presente estudio ha sido apoyado por subvenciones del Instituto de Salud Carlos IIIcofinanciado por el Fondo Europeo de Desarrollo Regional - FEDER (números decontrato: PI12 / 00378, SAS-PI-0239/2012, AC-0073-2013).
Conflicto de interesesLos autores declaran no tener ningún conflicto de intereses.
