
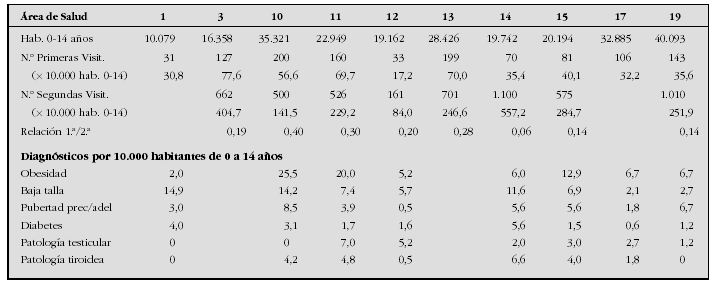
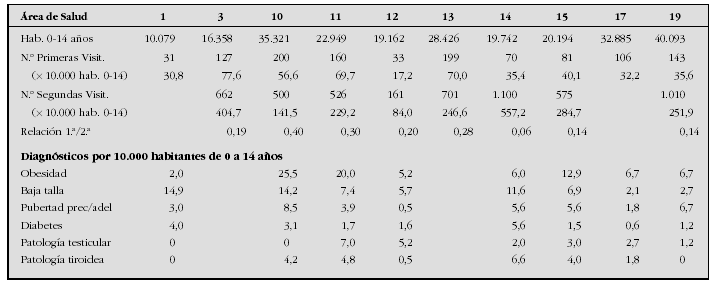
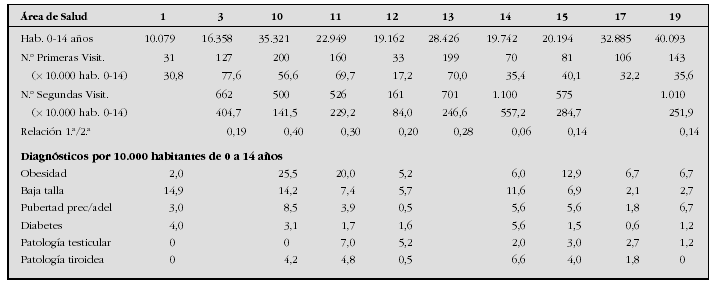
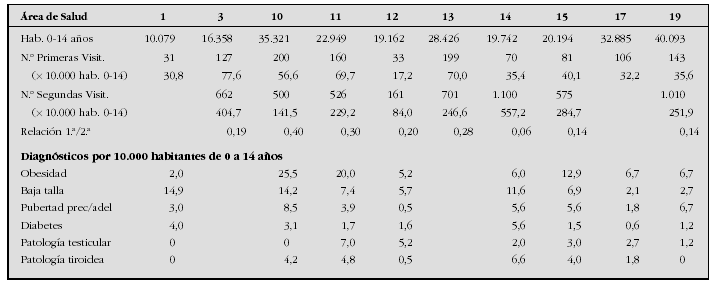
Introduction
Many metabolic pathways affect growth and changes in any gene in any of the component pathways can lead to different phenotypes due to decreased but in some cases to over expression of the perturbed gene. Selected disorders will be reviewed along with their molecular basis. Information on these disorders is found in Online Mendelian Inheritance in Man (OMIM) which is available at http://www.ncbi.nlm.nih.gov/Omim1. OMIM entry numbers corresponding to disorders in this chapter are given in the text and in table 1.

Growth hormone pathway
The growth hormone (GH) pathway is comprised of a series of interdependent genes whose products are required for normal growth (see fig. 1). The GH pathway genes include ligands (GH, insulin like growth factor 1 (IGF1), transcription factors (Prophet of PIT1 or PROP1 and PIT1), agonists (GH releasing hormone or GHRH), antagonists (somatostatin or GH1F) and receptors (GH releasing hormone receptor or GHRHR and the GH receptor or GHR). These genes are expressed in different organs and tissues including the hypothalamus, pituitary, liver and bone (see fig. 1). The effective and regulated expression of the GH pathway is essential for growth in stature as well as homeostasis of carbohydrate, protein and fat metabolism. Figure 1 shows the GH pathway including its' interdependent genes that are required to produce transcription factors, ligands such as GH, receptors such as GHR and other gene products needed for normal growth. Known gene defects in the GH pathway that cause disease including the following: 1) CPHD - HESX1, LHX3, PROP1 and PIT1; 2) IGHD - GH1 and GHRHR and 3) GH Resistance - GHR and IGF1.

Figure 1. The Growth Hormone (GH) biosynthetic pathway (for details see text).
Isolated growth hormone deficiency (IGHD)
Estimates of the frequency of GH deficiency (GHD) range from 1/4,000 to 1/10,000 in various studies. Causes of GHD include central nervous system insults or defects such as cerebral edema, chromosome anomalies, histiocytosis, infections, radiation, septo-optic dysplasia, and trauma or tumors that affect the hypothalamus or pituitary. While most GHD cases are the only member of their family, estimates of the proportion of cases that have an affected parent, sib or child range from 3 to 30 % in different studies. This familial clustering suggests that having a close affected relative conveys substantial relative risk and that a significant proportion of GHD cases may have a genetic basis2-3. Our current understanding of the genetic contributions to a series of familial defects in the GH pathway will be discussed in the following sections.
Clinical features
An intact GH pathway is needed throughout childhood to maintain normal growth. Concomitant or combined deficiencies of other pituitary hormones (LH, FSH, TSH, and/or ACTH), in addition to GH, is called combined pituitary hormone deficiency (CPHD) or panhypopituitary dwarfism. The combination of GH and these additional hormone deficiencies often causes more severe retardation of growth and skeletal maturation and spontaneous puberty may not occur2-3.
Diagnosis
Short stature, delayed growth and delayed skeletal maturation occurs with GH pathway defects. Since these signs can also be associated with systemic illnesses, individuals suspected to have GHD should be evaluated for systemic diseases before having complicated tests to detect GHD. Provocative tests for GHD include GH stimulation tests. Deficient GH peak responses range from 7-10 ng/ml. Testing for concomitant deficiencies of LH, FSH, TSH, and/or ACTH should be done on GHD cases to detect CPHD to provide a complete diagnosis and enable planning of optimal treatment2-3.
Types of familial IGHD
That are associated with at least six different Mendelian disorders. These include four autosomal recessive disorders (IGHD 1A and 1B, Bioinactive GH and GHRHR defects) (OMIM#s 262400, 139250, 262650 and 139191, respectively). In addition, there is an autosomal dominant (IGHD II, OMIM 173100) and an X-linked form of IGHD (IGHD III, OMIM# 307200)(see table 1).
IGHD IA: The most severe form of IGHD, called IGHD IA, (OMIM 262400 and139250) has an autosomal recessive mode of inheritance. Affected neonates occasionally have mildly decreased birth lengths and hypoglycemia in infancy. All develop severe dwarfism by 6 months of age. While replacement therapy with exogenous GH gives a good initial growth response in individuals with IGHD IA, this response is often temporary because GH resistance develops due to anti-GH antibodies. IGHD IA is usually caused by a deletion of the GH1 genes. At a molecular level, these DNA deletions are 6.7, 7.0 or 7.6 kb in length with ~3/4 being 6.7 kb2-3. DNA sequence analysis of the fusion fragments associated with these recurring deletions has shown that the deletions arise from homologous recombination between repeated sequences that flank the GH1 gene4. Multiple studies indicate that ~15 % of individuals with severe IGHD (> 4.5 SD in height) have GH1 gene deletions. Since gene deletions, as well as frameshift and nonsense mutations have been found to cause the IGHD IA phenotype, this disorder is best described as complete GHD due to heterogeneous GH1 gene defects, rather than gene deletions alone (see table 1)2-3.
IGHD IB: This milder form of IGHD also has an autosomal recessive mode of inheritance. IGHD IB These cases differ clinically from IGHD IA in their having low but detectable levels of GH and a continued growth response due to immunological tolerance to treatment with exogenous GH. IGHD IB cases are caused by GH gene defects that result in a mutant GH protein that may not be detected by RIA. The presence of these mutant GH protein molecules may explain the good responses that are seen to GH therapy because their presence mitigates against the production of anti-GH antibodies. IGHD 1B is caused by mutations that affect splicing of the GH1 gene. This altered splicing causes loss of amino acids that affect the stability and biological activity and reduce secretion of the mutant GH protein2-3.
IGHD II: IGHD II has an autosomal dominant mode of inheritance due to dominant-negative mutations of the GH1 gene and respond well to GH treatment. Almost all the GH1 gene defects reported in IGHD II are mutations that alter splicing of GH mRNA and cause skipping or deletion of exon 3. The mechanism by which these dominant negative mutations prevent expression of GH protein from the other, normal GH1 gene is poorly understood. Other IGHD II mutations cause skipping of exon 3 by disrupting splicing enhancer sequences (Ses) that regulate the splicing pattern of GH mRNA and, when these Ses are perturbed, exon 3 skipping occurs2-3. An IGHD II mutation that does not cause abnormal splicing is a G to A transition which results in an Arg to His substitution at residue 183 (Arg183His) of the GH molecule. This substitution is thought to alter the intracellular processing of the GH molecule by binding to zinc, thereby deranging the zinc associated presecretory packaging of GH5.
IGHD III: A third form of IGHD called IGHD III (OMIM 307200) has an X-linked mode of inheritance, and distinct clinical findings in different families. In some families all cases have agammaglobulinemia associated with their IGHD, while in other families all cases have only IGHD. This suggests that contiguous gene defects on the long arm of the x chromosome may cause some IGHD III cases. Duriez et al reported that X-linked agammaglobulinemia and IGHD is caused by mutation in the Bruton's tyrosine kinase or BTK gene6.
Laumonnier et al studied the SOX3 gene in families with X-linked mental retardation where the causative gene had been mapped to Xq26-q277. They showed that the SOX3 gene maps to Xq26.3 and was involved in a large family in which affected individuals had mental retardation and IGHD (OMIM 300123 and 313430, table 1). The mutation was an in-frame duplication of 33 bp encoding 11 alanines in a polyalanine tract of the SOX3 gene. The expression pattern during neural and pituitary development suggested that dysfunction of the SOX3 gene caused by this polyalanine expansion might disturb transcription pathways and the regulation of genes involved in pituitary development.
Biodefective GH
A number of patients have been described with the clinical features of IGHD who achieved normal plasma immunoactive GH levels following GH provocative or stimulation tests, but low levels of somatomedin (OMIM 139250, table 1). Less GH was detected by radioreceptor assay than by RIA analysis in some studies. In view of their clinical syndrome of IGHD, apparently normal plasma concentrations of GH, low basal somatomedin levels, and their normal responce to exogenous GH, individualos with Bioinactive GH are thought to secrete a biologically inert GH. Takahashi et al identified a C to T transition in codon 77 which results in an Arg to Cys substitution in the GH1 gene of a subject diagnosed with bioinactive GH8.
GHRH receptor defects
A variety of mutations have been detected in the human GHRHR gene in individuals with IGHD (OMIM 139191). In a kindred with a nonsense mutation affected family members had poor growth since infancy and were extremely short. They failed to produce GH in response to standard provocative tests and had good responses to GH replacement. Cases were homozygous for a G to T transversion that caused a premature termination mutation (Glu72Stop). A large Brazilian family was reported by Salvatori et al who had many family members with IGHD due to an intronic G to T transition that destroys the 5' splice site of IVS 1 of the GHRHR gene9.
Growth hormone resistance
To be biologically active, GH (OMIM 139250) must bind to a transmembrane receptor (GHR) (OMIM 60094), the GHR must form a dimer, and an intracellular signal-transduction pathway must be activated which causes the synthesis and secretion of insulin-like growth factor I (IGF1) (OMIM 147440). This factor, which in serum is bound to members of a family of binding proteins, binds to the IGF1 receptor (IGF1R) (OMIM 147370) and activates its own signal-transduction pathways, resulting in mitogenic and anabolic responses that lead to growth. It is uncertain whether GH has any anabolic actions independent of IGF1. Disruptions in GHR or IGF1 can cause GH resistance characterized by phenotypic features of GH deficiency associated with normal or high GH levels.
Growth hormone receptor dysfunction or laron dwarfism I
This an autosomal recessive disorder caused by GH resistance due to defects in the GH receptor (GHR) gene (OMIM 262500). While at the clinical level Laron syndrome cases are indistinguishable from GHD cases they differ at the biochemical level because they have low levels of IGF1, despite their having normal or increased levels of GH (see fig. 1). This contrasts with the low levels of both IGFI1 and GH that are seen in GHD. Importantly, exogenous GH does not induce an IGF1 response or restore normal growth in Laron dwarfism I cases because their GHR dysfunction prevents the synthesis and secretion of IGF1. While plasma levels of the GH binding proteins (GHBP) that are derived from the extracellular domain of GHR, are usually low in Laron Dwarfism I cases, Woods et al reported a homozygous point mutation in the intracellular domain of the GHR that caused Laron syndrome with elevated GHBP levels10. These authors predicted that the mutant GHR would not be anchored in the cell membrane but would be measurable in the serum as GHBP, thus explaining the phenotype of severe GH resistance combined with elevated circulating GHBP. Studies of the GHR genes of Laron Dwarfism I cases (OMIM 262500) have identified a variety of exon deletions and base substitutions. While treatment with exogenous GH is ineffective in those with GHR dysfunction, replacement therapy with recombinant IGFI1 has been shown to be effective (see fig. 1 and table 1).
Laron dwarfism II
This form of GH resistance is caused by post GHR defects (see OMIM 245590, table 1). Patients with Laron dwarfism II have elevated serum GH, normal GHBP levels, and respond well to treatment with IGF1 indicating that their growth deficiency is due to a post-GHR defect (see fig. 1). Woods et al described a patient with severe growth failure, sensorineural deafness, and mental retardation who was homozygous for a partial deletion of the IGF1 gene11.
Combined pituitary hormone deficiency (CPHD)
Cases with combined pituitary hormone deficiency (CPHD) vary in their clinical findings because they have deficiencies of varying severity of one or more of the other pituitary trophic hormones (ACTH, FSH, LH or TSH) in addition to GHD (OMIM 262600). While most cases of CPHD are sporadic, a variety of familial forms are known that can have autosomal recessive, autosomal dominant, or X-linked modes of inheritance.
HESX1 mutations
HESX1 is expressed in the thickened layer of oral ectoderm that gives rise to the Rathke pouch, the primordium of the anterior pituitary. Down regulation of HESX1 coincides with the differentiation of pituitary-specific cell types. Dattani et al found a missense HESX1 mutation (ARG53CYS) in homozygous state in a brother and sister with septooptic dysplasia, agenesis of the corpus callosum and CPHD (OMIM 182230)12.
LHX3 mutations
Murine Lhx3 mRNA accumulates in Rathke's pouch, the primordium of the pituitary and may be involved differentiation of pituitary cells. Netchine et al identified two families with CPHD (OMIM 262600) caused by mutations in the LHX3 gene13. The phenotype associated with these mutations included the following: 1) severe growth retardation, 2) complete deficiency of all but one of the anterior pituitary hormones (ACTH), 3) elevated and anteverted shoulders with a short neck associated with severe restriction of rotation of the cervical spine and 4) an enlarged anterior pituitary. The authors concluded that LHX3 is required for the proper development of all anterior pituitary cell types except corticotropes and that the rigid cervical spine phenotype is consistent with a function of LHX3 in the proper development of extrapituitary structures as well.
PIT1 mutations
Defects in the PIT1 gene causes familial CPHD cases which have a different phenotype (OMIM 173110). PIT1 is an anterior pituitary-specific transcription factor, which regulates the expression of GH, PrL and TSH. PIT1 is also required for pituitary cellular differentiation and function. PIT1 has functional domains that enable transactivation of other genes including GH, PrL and TSH or binding to these genes. At least six different PIT1 mutations causing autosomal recessive and two others causing autosomal dominant CPHD have been found in humans in a subtype of panhypopituitary dwarfism associated with GH, PrL and TSH deficiency (see table 1).
PROP1 mutations
PROP1 or Prophet of PIT1 is a pituitary specific homeodomain factor that is required for development of somatotropes, lactotropes, thyrotropes of the anterior pituitary and for expression of PIT1. Multiple PROP1 gene mutations cause an autosomal recessive CPHD that has a third phenotype in humans (OMIM 601538, table 1). In addition to deficiencies of GH, PrL, and TSH that are seen in those with PIT1 defects, subjects with PROP1 defects also have deficiencies of LH and FSH, which prevent the onset of spontaneous puberty and, in some cases, ACTH deficiency in later life. The various PROP1 mutations include 1) a C to T transition in codon 120 which encoded a TGC (Arg) to CGC (Cys) substitution, 2) a T to A transversion that encodes a TTC (Phe) to ATC (Ile) substitution at codon 117 and 3) 2 bp AG deletion in codon 101 (101delAG) that causes a frameshift and results in a premature stop at codon 109. The resulting protein products from all three of these different PROP1 mutations have greatly reduced DNA binding and transactivation abilities14. The 101delAG is a recurring mutation that is estimated to occur in about 55 % of familial and 12 % of sporadic CPHD cases15. A fourth PROP1 mutation is a 2 bp GA deletion in codon 51 (51delGA)16. Like the 101delAG mutation, the 51delGA mutation causes a frameshift that results in a premature stop codon. This mutation was found in 12 % of familial and 21 % of sporadic CPHD cases.
X Linked CPHD
Lagerstrom-Fermer et al reported a family, which included affected males suffering from variable degrees of CPHD (OMIM 312000)17. Some affected males who died during the first day of life and had postmortem findings of hypoadrenalism, presumed to be due to CPHD. Others had variable combinations of hypothyroidism, delayed pubertal development, and short stature due to GHD. All surviving patients exhibited mild to moderate mental retardation. They found linkage with markers in the Xq25-q26 region. Furthermore, they found an apparent extra copy of the marker DXS102 in affected males and heterozygous carrier females, suggested that a segment including this marker was duplicated.
Treatment of pituitary gene mutations
Recombinant derived GH is widely available but must be given by subcutaneous injection. To obtain an optimal outcome, children with GHD should be started on replacement therapy as soon as their diagnosis is established. The dosage increases with increasing body weight to a maximum during puberty and is usually discontinued by ~17 years of age. Disorders in which GH treatment is of proven efficacy include GHD, either isolated or in association with CPHD and Turner syndrome. The clinical responses of individuals with IGHD or CPHD to GH replacement therapy varies depending on 1) the severity and age at which treatment is begun, 2) recognition and response to treatment of associated deficiencies such as thyroid hormone deficiency and 3) if treatment is complicated by the development of anti-GH antibodies. The outcome of Turner syndrome subjects varies with the severity of their 1) short stature, 2) chromosomal complement and 3) age when treatment began3.
Disorders of hyper secretion and/or neoplasia
Genetic disorders of pituitary hyperfunction are far less common than those of pituitary insufficiency (see table 1). Furthermore, except in instances of multiple endocrine neoplasia (MEN) or in patients who have a positive family history of pituitary hyperfunction, it is difficult to determine which of the sporadic cases have a genetic form of pituitary disease. The most common form of hereditary pituitary neoplasia is MEN, type I. Although multiple cases of familial acromegaly and of the amenorrhea-galactorrhoea syndrome have been described in certain kindreds with no evidence of other endocrine involvement, these disorders may well represent limited forms of MEN.
Acromegaly
Hyper secretion of GH by a pituitary neoplasm leads to the classic syndromes of acromegaly and gigantism, depending upon the age of onset of the disorder (see OMIM 102200, table 1). The diagnosis can be made readily on the basis of high fasting plasma concentrations of GH, which usually do not fall following oral glucose ingestion. Although the majority of cases of acromegaly are sporadic, many families have been reported in which multiple members were thought to be affected. Another syndrome that might be confused with acromegaly is cerebral gigantism, in which increased growth rate occurs from infancy but no abnormalities in GH secretion are found. In spite of these reservations, pathological and radiographic documentation of pituitary adenomas have been described in successive generations in several families, strongly suggesting autosomal dominant inheritance.
Activating mutations of G protein and subunit genes
While in most cases defects in the GH pathway cause deficient growth, there are defects that can cause excess growth. Included among these are cerebral gigantism, acromegaly and McCune-Albright or Albright Hereditary Osteodystrophy syndrome. Pathway defects causing cerebral gigantism and its adult counter part, acromegaly, include somatic mutations in a G protein ? subunit GS? (GNAS) gene (OMIM 139320 and 174800). These de novo mutations in GS? inhibit GTPase activity and cause inappropriate and continuous stimulation of adenylyl cyclase (see fig. 2). If such mutations occur in the pituitary they can cause adenomas that autonomously secrete inappropriately large amounts of GH despite the lack of GHRH stimulation and the presence of somatostatin inhibition (see fig. 1). McCune-Albright syndrome (OMIM 174800) is a sporadic growth disorder, which has the clinical findings of polyostotic fibrous dysplasia, café au lait spots, sexual precocity, and endocrine disorders of over production. In some cases the somatic GS? gene mutations causing McCune-Albright syndrome can activate unregulated GH production by affected somatotropes18.

Figure 2. Mutations affecting the cyclic AMP pathway (left) can cause McCune Albright syndrome.B12-12.
Cytochrome P 450 genes
These genes are important in many metabolic pathways. Their contribution to growth disturbances include the congenital adrenal hyperplasias due to mutations in CYP21, 11A and 11B1 and STAR, as well as height variations that associate with polymorphisms in CYP19.
Congenital adrenal hyperplasia (CAH)
CAH can be caused by deficiencies of one of a variety of the enzymes in the cortisol biosynthesis pathway (see OMIMs 201910, 600617, 201710 and 202010). CAH affects about 1 in 5,000 births and in about 95 % of cases, 21-hydroxylation is impaired (see OMIM 201910) in the adrenal cortex so that 17-hydroxyprogesterone (17-OHP) is not converted to 11-deoxycortisol. Because of defective cortisol synthesis, ACTH levels increase, resulting in overproduction and accumulation of cortisol precursors, particularly 17-OHP, proximal to the block. This causes excessive production of androgens, resulting in virilization. In female newborns, the external genitalia are masculinized; gonads and internal genitalia are normal. Postnatally, untreated males as well as females may manifest rapid growth, penile or clitoral enlargement, precocious adrenarche, and ultimately early epiphyseal closure and short stature. A mild form of late-onset adrenal hyperplasia due to 21-hydroxylase deficiency can occur in adults and has hirsutism as the only manifestation in the most attenuated phenotype. CAH can be caused by heterogeneous CYP21 mutations including deletions, frameshifts, missense, nonsense and slicing defects. CAH is caused by homozygosity or compound heterozygosity for these various CYP21 alleles.
Aromatase (CYP19)
Aromatase or estrogen synthetase, catalyzes the formation of aromatic C18 estrogens from C19 androgens (OMIM 107910). Aromatase is present in many tissues including skin, muscle, fat, and nerve, where it may contribute to sex-specific differences in cellular metabolism. Ellis et al performed an association study using common polymorphisms in CYP19 and the Y chromosome19. They detected an association between CYP19 and height, that was more evident in men than in women. An association was also found with the Y chromosome. When men were grouped according to haplotypes of the CYP19 and Y chromosome polymorphisms, a difference of 4.2 cm was detected. The authors concluded that in men, genetic variation in CYP19 and on the Y chromosome are involved in determining normal adult height, and that these loci may interact in an additive fashion.
Mendelian disorders with endocrine abnormalities
A variety of Mendelian disorders have among their pleiotropic effects endocrine abnormalities. These disorders include some, such as achondroplasia, that have a single common mutation and others, such as hemoglobinopathies, that are caused by heterogeneous mutations.
Achondroplasia with obstructive sleep apnea
Achondroplasia is a common skeletal dysplasia in which the dwarfism is due to an abnormality in endochondral ossification (see OMIM 100800, table 1). Up to 10 % of patients with achondroplasia have been reported to have serious respiratory complications. Goldstein et al studied a 9-year-old boy with achondroplasia and obstructive sleep apnea who had low-growth hormone secretion during sleep (20). Following tracheostomy his GH secretion during sleep normalized, and his growth rate almost doubled. This case suggests that short patients with craniofacial syndromes associated with obstructive sleep apnea may have diminished sleep entrained GH secretion as a contributing cause to their growth retardation.
Borjeson-Forssman-Lehmann syndrome
The X-linked Borjeson-Forssman-Lehmann syndrome is characterized by short stature, hypogonadism, hypotonia, severe mental deficiency, coarse facial appearance with a prominent brow ridge and large ears in affected males (see OMIM 301900, table 1). Robinson et al documented markedly deficient GH responses to arginine and L-dopa, as well as low somatomedin C levels in a severely affected male and two of his mildly affected twin sisters21. The growth deceleration in this syndrome, however, may not begin until age 8-10 years, and therefore the pituitary deficiency may be progressive. Indeed, the associated hypogonadism has been reported to be both primary and secondary, and may also evolve with age.
CHARGE Association
CHARGE is an acronym that describes a nonrandom association of anomalies: colobomas of the eye; heart disease; atresia of the choanae; retarded growth, development and/or CNS anomalies; genital hypoplasia; and ear anomalies or deafness (see OMIM 214800, table 1). Growth retardation, which is usually of postnatal onset, and hypogonadism are prominent features of the CHARGE syndrome and may well be due to hypothalamic defects. August et al documented GH and gonadotropin deficiencies in a girl with the CHARGE association, who also had a delayed peak TSH response to TRH22. Boys may have cryptorchidism, microphallus and/or hypospadias. Adult males were also reported to have genital hypoplasia and no secondary sexual characteristics. Arrhinencephaly and holoprosencephaly have been reported, suggesting a hypothalamic cause for the pituitary deficiencies. Although most cases are sporadic, a few familial occurrences suggestive of both dominant and recessive inheritance have been reported.
Fanconi anemia
The Fanconi syndrome is an autosomal recessive disorder characterized by chronic pancytopenia with bone marrow hypoplasia, abnormal pigmentation, upper limb malformations, kidney anomalies, growth retardation, small genitalia, and increased frequency of chromosomal breaks in cultured lymphocytes (see OMIMs 227650, 227660, 227645, 227646 and 600901 table 1). Nilsson found that 38/68 (56 %) of published cases of Fanconi anemia had stunted growth and 24/68 (35 %) had genital anomalies23. Cussen reported a child with Fanconi anemia who appeared to be a pituitary dwarf and pointed out that small pituitary glands, adrenocortical atrophy and atrophic testis have been described in this syndrome24. A number of investigators have now documented GH deficiency in patients with Fanconi anemia, in most of whom other endocrine function was normal. Administration of GH resulted in excellent short-term and long-term responses in most of these patients. In view of the intrauterine growth retardation commonly associated with this syndrome, it appears that both cellular factors and GH deficiency probably contribute to the short stature.
Hemochromatosis
Male hypogonadism and pituitary hemosiderosis can both occur in hemochromatosis (see OMIMs 235200 and 602390, table 1) and abnormalities have also been found in gonadotropin, cortisol, GH, PrL and TSH secretion. Stocks and Martin found that functional pituitary insufficiency of varying degree occurs in 60 % of patients with hemochromatosis25. Signs and symptoms of gonadal dysfunction included depressed sexual function, testicular atrophy, absent urinary gonadotropins, decreased plasma levels of LH, and low plasma testosterone levels, indicating that the hypogonadism in hemochromatosis is secondary to a deficiency of pituitary gonadotropin. The testes of hypogonadal hemochromatosis patients usually show evidence of secondary atrophy without iron deposition, documenting the hypogonadotropic nature of the hypogonadism in this disease. Most studies have reported testicular atrophy with low levels of gonadotropins, and unresponsive to gonadotropin-releasing hormone. Hemochromatosis is an autosomal recessive trait tightly linked to the HLA loci.
Hemoglobinopathies
There are well-documented cases of acquired pituitary insufficiency occurring in adults with hemoglobinopathies, presumably secondary to infarction of the gland (see OMIM 141900, table 1). In one instance, a 41-year-old American black person with sickle-cell trait (hemoglobin SA) developed fatigue, weight loss, decreased libido, impotence, polyuria and polydypsia a few months after a prolonged high altitude flight26. Endocrinologic evaluation revealed evidence of both anterior and posterior pituitary insufficiency. Among 130 autopsied cases of sickle trait (SA), McCormick found two instances of pituitary infarction and abnormal GH secretion has been found thalassemic patients on clinical transfusion therapy27.
Histiocytosis x (Letterer-Siwe Disease, Hand-Schuller-Christian Disease, Eosinophilic Granuloma)
Histiocytosis x is characterized by foamy histiocyte infiltration in many areas of the body, including the hypothalamus. When the histiocytic infiltration involves the hypothalamus, prepubertal growth retardation associated with GH deficiency and diabetes insipidus frequently occur (see OMIM 246400, table 1). Delayed puberty and hypogonadism are also frequent accompaniments of this syndrome. Autopsy reports in adults with histiocytosis x suggest that the pituitary insufficiency is secondary to hypothalamic destruction. Diabetes Insipidus and GHD frequently occur together, but either endocrine abnormality may exist alone. In contrast to a previous suggestion of GH unresponsiveness, Braunstein et al documented a significant increment in growth rate in response to GH therapy in these individuals28.
Neurofibromatosis type 1
A variety of endocrine disturbances has been reported in patients with neurofibromatosis which has an autosomal dominant mode of inheritance (see OMIM 162200, table 1). The most common associated endocrine disorder in children with NF1 is sexual precocity, while pheochromocytoma is the most common in adults. Marked growth retardation, unrelated to skeletal anomalies, has also been reported. Andler et al documented a variety of pituitary dysfunctions in affected children, including GH deficiency, both diminished and elevated TSH response to TRH, and hyperprolactinemia29. All of their patients with neurofibromatosis and pituitary dysfunction had a suprasellar tumor.
Pallister-Hall Syndrome (Congenital Hypothalamic Hamartoblastoma, Hypopituitarism, Imperforate Anus, Postaxial Polydactyly Syndrome)
Hall et al described this neonatally lethal malformation syndrome which consists of hypothalamic hamartoblastoma, hypopituitarism, postaxial polydactyly and imperforate anus (see OMIM 146510, table 1)30. Variable features include laryngeal cleft, abnormal lung lobulation, renal agenesis and/or renal dysplasia, short 4th metacarpals, nail dysplasia, multiple buccal frenula, hypoadrenalism, microphallus, congenital heart defect and intrauterine growth retardation. An anterior pituitary gland was absent in all cases. The posterior pituitary was absent in the majority. The adrenal hypoplasia, small thyroid and microphallus are presumably secondary to pituitary insufficiency. Most cases have been sporadic, but several instances of father to son transmission have been reported, suggesting autosomal dominant inheritance.
Rieger Syndrome (Iris-Dental Dysplasia)
The Rieger syndrome is an autosomal dominant disorder associated with malformation of the iris, with pupillary anomalies and hypoplasia of the teeth, with or without maxillary hypoplasia. Sadeghi-Nejad and Senior reported a large family in which multiple individuals had both Rieger syndrome and IGHD (see OMIM 180500, table 61-1)31. Sibs of the proband had Rieger syndrome with normal pituitary function, but GHD was not found in any member of the family who did not have Rieger syndrome. Affected individuals had insulin hypersensitivity, but normal plasma insulin responses to arginine and glucose. One subject who was treated with GH exhibited substantial enhancement of his rate of growth. It is postulated that the basic pathogenetic mechanism in this autosomal dominant disorder is maldevelopment of the neural crest, resulting in ocular, dental and hypothalamic abnormalities. Primary empty sella with normal pituitary function has also been reported in association with dominantly inherited Rieger syndrome in multiple members of a large kindred.
Chromosomal disorders with endocrine abnormalities
A large variety of chromosomal abnormalities are associated with endocrine disorders. These chromosomal anomalies can affect a variety of autosomes as well as the sex chromosomes.
Autosomal
GH deficiency has been described with 18p- and 20p- chromosomal deletions (see OMIM 146390, table 2)32-33. In addition, molecular detection of deletions of 17q (i.e., the GH gene cluster region) has been documented in IGHD IA. Since the gene for GHRH has been mapped to 20p, the GH deficiency in 20p- could result from either the deletion of the GHRH gene or a developmental anomaly of the hypothalamus. Patients with 18p- has been reported with hypopituitarism and solitary central maxillary incisor, suggesting that the pituitary insufficiency in 18p- may be due to a structural malformation of the hypothalamus. GHD has been rarely associated with 47XXY, 49XXXXY, and ring 5. Finally a search of the London Dysmorphology Database at http://www.hgmp.mrc.ac.uk/DHMHD/view.html identifies a series of partial chromosome deletions or duplications that are associated with short stature and pituitary abnormalities (see table 2). These include the following deletions: del(4)pter-p16, del(7)q32-qter, del(13)q22-qter, del(14)q22-q23, del(18)p, del(18)q21-qter, del(22)pter-q11 and duplications: dup(1)q25-q32, dup(9)p, dup(9)pter-q22 and dup(11)q23-qter.

Turner syndrome (45, X)
Although GH secretion has been reported to be normal, or paradoxically increased, in most patients with gonadal dysgenesis, pituitary insufficiency has been reported in several patients (see table 2). Ross et al studied GH secretion in 30 patients with Turner syndrome and found no differences in mean GH concentration or peak amplitudes throughout the day and night between patients less than 8 years of age and controls34. Patients over 9 years of age had lower mean GH levels and peak amplitudes. Reduced plasma somatomedin C levels and delayed bone age were found in patients of all ages. These abnormalities in GH secretion in Turner syndrome are probably secondary to the absence of sex hormones during adolescence.
Thyroid hormone abnormalities
Genetic defects in thyroid hormone biosynthesis include mutations in the thyroid-stimulating hormone (TSHB), thyroid-stimulating hormone receptor (TSHR) as well as organification defects among others. In this section selected defects will be reviewed.
Thyroid-releasing hormone (TRH) deficiency
TRH is the major hypothalamic mediator of release of thyroid-stimulating hormone (TSH). TRH is a tripeptide (glu-his-pro) that stimulates the release of both TSH and prolactin. The prolactin responce is enhanced in hypothyroidism and diminished in hyperthyroidism. Niimi et al reported a girl with isolated TRH deficiency presumed to be due to an autosomal recessive defect (see OMIM 275120)35.
Thyroid-releasing hormone receptor (TRHR) deficiency
Isolated central hypothyroidism, due to deficient TSH secretion causes low levels of thyroid hormones. Collu et al reported a 9 year old boy who had complete absence of TSH and PRL responses to TRH36. They detected a 9-bp deletion of nucleotides 343 to 351of the TRHR gene, leading to the deletion of amino acid residues 115-117 (Ser-Ile-Thr) and a G to A transition encoding an Ala118Thr substitution. The maternal allele had a C to T transition resulting in an Arg17Ter mutation (OMIM 188545). Mutated TSH receptors were unable to bind TRH and to activate total inositol phosphate accumulation and the authors concluded that they were naturally occurring inactivating mutations of a G protein-coupled receptor linked to the phospholipase C second-messenger pathway (see fig. 2).
Thyroid-stimulating hormone (TSH) deficiency
TSH is also called thyrotropin. In a family with 2 affected sisters born of consanguineous parents, Hayashizaki et al found homozygosity for a G to A transition in exon 2 in codon 29 of the TSH-beta subunit37. This transition encoded a Gly29Arg substitution in the center of the so-called CAGYC region, which represents an amino acid sequence conserved among all known glycoprotein hormone beta subunits (OMIM 188540).
Thyroid-stimulating hormone receptor (TSHR) deficiency
Trulzsch et al described a database of TSHR mutations that includes a summary of mutations identified and the variety of clinical phenotypes associated with the different mutations38. These include activating mutations in toxic thyroid nodules, germline mutations as the cause of sporadic and familial nonautoimmune autosomal dominant hyperthyroidism and inactivating mutations associated with inherited TSH insensitivity or hypothyroidism (see OMIM 603372).
Pendred syndrome (PDS)
PDS is an autosomal recessive disorder associated with developmental abnormalities of the cochlea, sensorineural hearing loss, and diffuse thyroid enlargement (goiter). PDS is caused by mutations in the SLC26A4 gene (see OMIM 605646).

